No more microevolution! No more evolution at all...
“I myself am convinced that the theory of evolution, especially the extent to which it’s been applied, will be one of the great jokes in the history books in the future. Posterity will marvel that so very flimsy and dubious an hypothesis could be accepted with the incredible credulity that it has.” - Malcolm Muggeridge
Another reason why the creationist position often appears as one of weakness is that we are perceived as admitting variation only because of being forced to do so by observation, then simply escaping the implications of variation by saying it does not go far enough. And we appear to redraw our ‘Maginot line’ depending on how much variation is demonstrated. It will be shown shortly, though, that this is a caricature of the creationist position, and that the limits to variation arise from basic informational considerations at the genetic level.
However, the argument is circular and thus vulnerable. Since creationists by definition regard all variation as ‘within the limits of the created kind’ (see for example the statement of belief of the Creation Research Society of the USA), how can we then use observations to prove that variation is within the limits of the kind? To put it another way—of course we have never observed variation ‘across the kind’, since whatever two varieties descend from a common source, they are regarded as the same kind. It is no wonder that evolutionists are keen to press us for an exact definition of the created kind, since only then does our claim of ‘variation is only within the kind’ become non-tautologous and scientifically falsifiable.
Circular reasoning does not invalidate the concept of created kinds, however. In the same way, natural selection is also only capable of a circular definition (those who survive are the fittest, and the fittest are the ones who survive), but it is nevertheless a logical, easily observable concept. All we are saying is that arguments which are inherently circular cannot be invoked as independent proof of the kinds.
When I claim that such independent proof may not be possible by the very nature of things, this statement is in no way a ‘cop out’. For instance, let us say we happened upon the remnants of an island which had exploded, leaving behind the debris of rocks, trees, sand, etc. It may be impossible in principle to reconstruct the original positions of the pieces in relation to each other before the explosion. This does not, however, mean that it is not possible to deduce with a great degree of confidence that the current state of the debris is consistent with that sort of an explosion which was recorded for us by eyewitness testimony, rather than arising by some other mechanism.
In like manner, we can show that the observations of the living world are highly consistent with the biblically described concept of original created kinds, and inconsistent with the idea of evolution. This is best done by focusing on the underlying genetic/informational basis of all biological change. This is more realistic and more revealing than focusing on the degree or extent of morphological change.
The issue is qualitative, not quantitative. It is not that the train has had insufficient time to go far enough—it is heading in the wrong direction. The limits to variation—observed or unobserved—will come about inevitably because gene pools run out of ‘functionally efficient’ genetic information (or ‘teleonomic’ information). A full understanding of this eliminates the image of the desperately backpedalling creationist, redrawing his line of last resistance depending on what new observations are made on the appearance of new varieties.
It also defuses the whole issue of ‘micro’ and ‘macro’ evolution. I believe it is better for creationists to avoid these confusing and misleading terms altogether. The word ‘evolution’ generally conveys the meaning of the sort of change which will ultimately be able to convert a protozoon into a man or a reptile into a bird, and so on. I hope to show that in terms of that sort of meaning, we do not see any evolution at all. By saying ‘we accept micro but not macroevolution’ we risk reinforcing the perception that the issue is about the amount of change, which it is not. It is about the type of change.
This is not merely petty semantics, but of real psychological and tactical significance. Of course one can say that ‘microevolution’ occurs when this word is defined in a certain fashion, but the impact of the word, the meaning it conveys, is such as to make it unwise to persevere with this unnecessary concessional statement. Microevolution, that is, a change, no matter how small, which is unequivocally the right sort of change to ultimately cause real, informationally ‘uphill’ change, has never been observed.
In any case, leading biologists are themselves now coming to the conclusion that ‘macroevolution’ is not just ‘microevolution’ [using their terminology] extended over time. In November 1980 a conference of some of the world’s leading evolutionary biologists, billed as ‘historic’, was held at the Chicago Field Museum of Natural History on the topic of ‘macroevolution’. Reporting on the conference in the journal Science, Roger Lewin wrote:
Imagine now the first population of living things on the evolutionist’s ‘primitive earth’. This so-called ‘simple cell’ would, of course, have a lot of genetic information, but vastly less than the information in only one of its present-day descendant gene pools, e.g., man. The evolutionist proposes that this ‘telegram’ has given rise to ‘encyclopedias’ of meaningful, useful genetic sentences. (See later for discussion of ‘meaning’ and ‘usefulness’ in a biological sense.) Thus he must account for the origin with time of these new and meaningful sentences. His only ultimate source for these is mutation.3
Going back to the analogy of the printed page, the information in a living creature’s genes is copied during reproduction, analogous to the way in which an automatic typewriter reproduces information over and over. A mutation is an accident, a mistake, a ‘typing error’. Although most such changes are acknowledged to be harmful or meaningless, evolutionists propose that occasionally one is useful in a particular environmental context and hence its possessor has a better chance of survival/reproduction. By looking now at the informational basis for other mechanisms of biological variation, it will be seen why these are not the source of new sentences and therefore why the evolutionist generally relies on mutation of one sort or another in his scheme of things.
This sort of variation can only occur if there is a storehouse of such sentences available to choose from. Natural (or artificial) selection can explain the survival of the fittest but not the arrival of the fittest, which is the real question. These Mendelian variations tell us nothing about how the genetic information in the present stock arose. Hence, it is not the sort of change required to demonstrate ‘upward’ evolution—there has been no addition of new and useful ‘sentences’. And this is in spite of the fact that it is possible to observe many new varieties in this way—even new species. If you define a species as a freely interbreeding natural unit, it is easy to see how new species could arise without any ‘uphill’ change. That is, without the addition of any new information coding for any new functional complexity. For example, mutation could introduce a defect which served as a genetic barrier, or simple physical differences, such as the sizes of Great Dane and Chihuahua, could make interbreeding impossible in nature.
It is a little surprising to still see the occasional creationist literature clinging to the concept that no new species have ever been observed. Even if this were true, and there is some suggestion that it has actually been observed, there are instances of ‘clines’ in field observations which make it virtually certain that two now-isolated (reproductively) species have arisen from the same ancestral gene pool. Yet the very same creationists who seem reluctant to make that sort of admission would be quite happy to agree with the rest of us that the various species within what may be regarded as the ‘dog’ kind, including perhaps wolves, foxes, jackals, coyotes and the domestic dog, have arisen from a single ancestral kind. So why may this no longer be permitted to be happening under present-day observations? It is not only biblically and scientifically unnecessary, but it sets up a ‘straw man’ in the sense that any definite observation of a new species arising is used as a further lever with which to criticize creationists.
What we see in the process of artificial selection or breeding giving rise to new varieties, is a thinning-out of the information in the parent stock, a reduction in the genetic potential for further variation. If you try and breed a Chihuahua from a Great Dane population or vice versa, you will find that your population lacks the necessary ‘sentences’. This is because, as each variety was selected out, the genes it carried were not representative of the entire gene pool.
What appeared to be a dramatic example of change with the appearance of apparently new traits thus turns out, when its genetic basis is understood, to be an overall downward movement in informational terms. The number of sentences carried by each subgroup is reduced thus making it less likely to survive future environmental changes. Extrapolating that sort of process forward in time does not lead to upwards evolution, but ultimately to extinction with the appearance of evermore-informationally-depleted populations.
At this point some will argue that the terms ‘useful’, ‘meaningful’, ‘functional’, etc. are misused. They claim that if some change gives survival value then by definition it has biological ‘meaning’ and usefulness. But this assumes that living systems do nothing but survive—when in fact they and their subsystems carry out projects and have specific functions. That is, they carry teleonomic information. This is one of the essential differences between living objects and non-living ones (apart from machines). These projects do not always give rise to survival/reproductive advantages—in fact, they may have very little to do with survival, but are carried out very efficiently. The Darwinian assumption is always made, of course, that at some time in the organism’s evolutionary history, the project had survival/reproductive value. (For example, the archer-fish with its highly-skilled ‘hobby’ of shooting down bugs which it does not require for survival at the present time.) However, since these are nontestable assumptions, it is legitimate to talk about genetic information in a teleonomic sense, in isolation from any possible survival value.
The gene pools of today carry vast quantities of information coding for the performance of projects and functions which do not exist in the theoretical ‘primeval cell’. Hence, in order to support protozoon-to-man evolution, one must be able to point to instances where mutation has added a new ‘sentence’ or gene coding for a new project or function. This is so regardless of one’s assumptions on the survival value of any project or function.
We do not know of a single mutation giving such an increase in functional complexity. Probabilistic considerations would seem to preclude this in any case, or at least make it an exceedingly rare event, far too rare to salvage evolution even over the assumed multibillion year time span.
To illustrate further—the molecule haemoglobin in man carries out its project of transporting and delivering oxygen in red cells in a functionally efficient manner. A gene or ‘sentence’ exists which codes for the production of haemoglobin. There is a known mutation (actually three separate ones, giving the same result) in which only one letter in the sentence has been accidentally replaced by another. If you inherit this change from both parents, you will be seriously ill with a disease called sickle cell anaemia and will not survive for very long. Yet evolutionists frequently use this as an example of a ‘beneficial mutation’. This is because if you inherit it from only one parent, your red cells will be affected, but not seriously enough to affect your survival—just enough to prevent the malaria parasite from using them as an effective host. Hence, you will be more immune to malaria and better able to survive in malaria-infested areas. This shows us how a functionally efficient haemoglobin molecule became a functionally crippled haemoglobin molecule. The mutation-caused gene for this disease is maintained at high levels in malaria-endemic regions by this incidental phenomenon of heterozygote superiority. Its damaging effect in a proportion of offspring is balanced by the protection it gives against malaria. It is decidedly not an ‘upward’ change. We have not seen a new, efficient oxygen transport mechanism or its beginnings evolve. We have not seen the haemoglobin transport mechanism improved.
One more loose but possibly useful analogy. Let us say an undercover agent is engaged in sending a daily reassuring telegram from enemy territory. The text says ‘the enemy is not attacking today’. One day an accident occurs in transmission and the word ‘not’ is lost. This is very likely going to be a harmful change, perhaps even triggering a nuclear war by mistake. But perhaps, in a freak situation, it could turn out to be useful (for example, by testing the fail-safe mechanisms involved). But this does not mean that it is the sort of change required to begin to convert the telegram into an encyclopedia.
The very small number of ‘beneficial’ mutations actually observed are simply the wrong kind of change for evolution—we do not see the addition of new sentences which carry meaning and information. Again surprisingly, one often reads creationist works which insist that there is no such thing as a beneficial mutation. If benefit is defined purely in survival terms, then we would not expect this to be true in all instances, and in fact it is not—that is, there are indeed ‘beneficial’ mutations in that sense only.
Information depends on order, and since all of our observations and our understanding of entropy tells us that in a natural, spontaneous, unguided and unprogrammed process order will decrease, the same will be true of information. The physicist and communications engineer should not be surprised at the realisation that biological processes involve no increases in useful or functional (teleonomic) information and complexity. In fact, the net result of any biological process involving transmission of information (i.e., all hereditary variation) is conservation or loss of that genetic information.
This points back directly to the creation of the information, supernaturally, in the beginning. It is completely in harmony with the biblical concept of a world made ‘very good’ as a balanced, functioning whole, with decay only subsequent to the Fall. This is the reason why there are inevitable limits to variation, why the creationist does not have to worry about how many new ‘species’ the future may bring—because there is a limit to the amount of functionally efficient genetic information present, and natural processes such as mutation cannot add to this original storehouse.
Notice that since organisms were created to migrate out from a central point at least once and fill empty ecological niches, as well as having to cope with a decaying and changing environment, they would require considerable variation potential. Without this built-in genetic flexibility, most populations would not be present today. Hence the concept of biological change is in a sense predicted by the biblical model, not something forced upon it only because such change has occurred.
Take a hypothetical created kind A—truly a biological ‘species’ with perhaps a tremendous genetic potential. See Figure 1. (For the sake of simplicity, the diagram avoids the issue of what is meant by two of each kind aboard the Ark—however, the basic point is not affected.) Note that A may even continue as an unchanged group, as may any of the subgroups. Splitting off of daughter populations does not necessarily mean extinction of the parent population. In the case of man, the original group has not diverged sufficiently to produce new species.
Hence, D1, D2, D3, E1, E2, E3, P1, P2, Q1, Q2, Q3 and Q4 are all different species, reproductively isolated. But all the functionally efficient genetic information they contain was present in A. (They presumably carry some mutational defects as well).
Let us assume that the original kind A has become extinct, and also the populations X, B, C, D, E, P and Q. (But not D1, D2, etc.) If X carried some of the original information in A, which is not represented in B or C, then that information is lost forever. Hence, in spite of the fact that there are many ‘new species’ which were not originally present, we would have witnessed conservation of most of the information, loss of some, and nothing new added apart from mutations (harmful defects or just meaningless ‘noise’ in the genetic information). All of which is the wrong sort of informational change if one is trying to demonstrate protozoon-to-man evolution.
Classifications above species are more or less arbitrary groupings of convenience, based generally on similarities and differences of structure. It is conceivable that today, D1, D2 and D3 could be classified as species belonging to one genus, and E1, E2 and E3 as species in another genus, for example. It could also be that the groups B and C were sufficiently different such that their descendants would today be in different families. We begin to see some of the problems facing a creationist who tries to delineate today’s representatives of the created kinds.
Creatures may be classified in the same family, for example, on the basis of similarities due to common design while in fact they belong to two totally different created kinds. This should sound a note of caution against using morphology alone, as well as pointing out the potential folly of saying ‘in this case, the baramin is the family; in this case, it is the genus, etc.’ (Baramin is an accepted creationist term for ‘created kind’.)
There is no easy solution as yet to the problem of establishing each of these genetic relationships—in fact, we will probably never be able to know them all with certainty. Interbreeding, in vitro fertilization experiments, etc. may suggest membership of the same baramin but lack of such genetic compatibility does not prove that two groups are not in the same kind. (See earlier discussion—genetic barriers could arise via mutational deterioration.) However, newer insights, enabling us to make direct comparisons between species via DNA sequencing, open up an entirely new research horizon. (Although the question of where the funding for such extensive research will come from in an evolution-dominated society remains enigmatic.)
What then do we say to an evolutionist who understandably presses us for a definition of a created kind or identification of same today? I suggest the following for consideration:
Besides being a simple and obvious definition, it is axiomatic. Thus it is as unashamedly circular as a rolled-up armadillo and just as impregnable, deflecting attention, quite properly, to the real issue of genetic change.
The question is not—what is a baramin, is it a species, a family or a genus? Rather, the question is—which of today’s populations are related to each other by this form of common descent, and are thus of the same created kind? Notice that this is vastly removed from the evolutionist’s notion of common descent. As the creationist looks back in time along a line of descent, he sees an expansion of the gene pool. As the evolutionist does likewise, he sees a contraction.
As with all taxonomic questions, common sense will probably continue to play the greatest part. The Scriptures, the fossil record and common sense unite to prevent creationists doing too much ‘lumping together’ as we go back in time. For instance, it is conceivable (though not necessarily so) that crocodiles and alligators both descended from the same ancestral gene pool which contained all their functionally efficient genes, but not really conceivable that crocodiles, alligators and ostriches had a common ancestral pool which carried the genes for all three!
The article above has convinced me to give up the word "microevolution" and stick to "variation within kind", which is both more specific and less confusing. Baramin, by the way, comes from Bara (created) and Min (kind) so a Min is a kind but all Min were created at the beginning and the word from the Bible is Baramin, thus, Baraminology!
Baraminology is a scientific way to sort distinct kinds of animals and their subgroups. Linnaeus was attempting to do this with his classification system, which has been taken by Darwinists and twisted and sometimes laughably broken. There really is no word (family, genus, species) that works as well or is as specific as a baramin. All wolf-dog types are of one baramin, for instance.
"For instance, it is conceivable (though not necessarily so) that crocodiles and alligators both descended from the same ancestral gene pool which contained all their functionally efficient genes, but not really conceivable that crocodiles, alligators and ostriches had a common ancestral pool which carried the genes for all three!"
Two important quotes from the telling article from 2009 listed below:
(Concerning the British public's tendency to believe in Intelligent Design) "Or you may do so because you genuinely do not know of the evidence for evolution, have never had it explained to you, or because you just don’t understand it."
"It" being the hypothesis of evolution by mutations in combination with natural selection over long periods of time. Of course the evidence for Darwinism is all a matter of just-so stories, it is not even possible for natural selection to operate on organisms that do not yet exist and there is no means by which organisms can form from matter naturally. Darwin has no explanation for life or information so it is dead in the water from the beginning. It also runs in the wrong direction, as the article above illustrates.
“This is a good thing, because as evolution is arguably the most important concept underlying the life sciences,” he said, “providing children with an understanding of it [evolution] at the earliest possible age will surely help lay the foundations for a surer scientific understanding later on.”
Balderdash! Humbug! Tommyrot! Lies! Foolishness! Darwinism has been nothing but a hindrance to operational science. How many tonsils were removed when they might better have been kept?. How much medical knowledge was held back because researchers believed in the idea of vestigal organs? We now know of no vestigal structures in humans at all. Darwinism was and is a drag on science.
Today we have millions of dollars wasted scanning the skies for a message from ET and think of the wasted salaries of every single member of the NCSE, which is nothing more than a Darwinist censorship organization!
Researchers have to throw in an homage to Darwin somewhere early in their publications before then going into the nuts and bolts of research than do not utilize Darwin at all. If we took Darwin out of science, the research in real science would continue unabated. Only the people trying in vain to make life come from non-life, make apes appear to be people or furiously writing dumbed-down explanations for textbooks would be out of a job!
From the November/December Creation Matters Magazine:
Darwin Marketed to Kids (Great Britain)
There’s a move on to get Darwin’s ideas taught to kids. Britain is giving a “birthday present to Darwin,”
wrote Andrew Copson for The Guardian,1 in the form of a national curriculum for primary schools
that will mention evolution for the first time — and prohibit teaching of creationism or intelligent design
in science lessons.
The addition of evolution to elementary school curriculum was in response to a letter promoted by the
British Humanist Association and signed by “scientists and experts.”Copson was obviously delighted
with what he perceived as a long-overdue smackdown against intelligent design — a belief espoused
by the majority of his fellow Britons: Those who care about public reason are routinely shocked by opinion
polls and surveys showing high levels of credence given to the idea of intelligent design. The most
recent poll purported to demonstrate that a majority of Britons think that it should be taught alongside evolution
in schools.
To solve this problem, we have to know what causes it and there are two reasons why you might prefer
the idea of intelligent design to that of evolution. You may do so because your prior ideological convictions,
mostly to do with religious belief, simply don’t allow you to accept the evidence that is presented to you.
Or you may do so because you genuinely do not know of the evidence for evolution, have never had it
explained to you, or because you just don’t understand it. In a society as decreasingly religious as England,
it is impossible to believe that most of the people who do not accept evolution are motivated by ideology rather than ignorance. This means that the best way to solve the problem is through better education and that is
what makes the inclusion of evolution in the science curriculum as early as possible so important.
Repeatedly in his article, Copson characterized the Darwin doubters as ignorant, but refused to acknowledge any ideological bias on his part. He also made it seem as if all evidence is for evolution and against religion — despite numerous evidential claims by intelligent design against evolution. He spoke favorably of a slate of
new children’s books on evolution such as What Mr. Darwin Saw, How Whales Walked into the Sea, and Mammals Who Morph.
“This is a good thing, because as evolution is arguably the most important concept underlying the life sciences,” he said, “providing children with an understanding of it [evolution] at the earliest possible age will surely help
lay the foundations for a surer scientific understanding later on.”
A new company is making Darwin toys for tots. Charlie’s Playhouse2 offers “Evolution for kids” in the
form of apparel, games, cards, and a giant evolution timeline kids can hop and skip on. Of special interest
is their 30-second commercial, Why are we making evolution toys? The answer: nobody else is. The
commercial laments the thousands of toys about physics, biology, and chemistry — even all those popular dinosaur toys — that don’t mention evolution. “But we do!” a cartoony Darwin exclaims, dancing
proudly at the end of the video.
William Dembski, the double-PhD scholar of the intelligent design movement, has had enough with all this.
His latest article on Uncommon Descent3 is called “Getting over our love for Darwin.” In it he quotes
Malcolm Muggeridge, who wrote, “I myself am convinced that the theory of evolution, especially the extent to which it’s been applied, will be one of the great jokes in the history books in the future. Posterity will marvel that so very flimsy and dubious an hypothesis could be accepted with the incredible credulity that it has.”
Exactly
1. Copson, A. (2009, November 9). A birthday present
for Darwin. Guardian. Retrieved December
10, 2009, from
www.guardian.co.uk/commentisfree/belief/2009 /nov/09/evolution-primary-schools-science
2. Charlie’s Playhouse. 2009. Why are we making
evolution toys? Retrieved December 20, 2009,
from http://www.charliesplayhouse.com/whyevolution- toys.php
3. Dembski, W.A. (2009, November 3). Getting over
our love for Darwin. Uncommon Descent. Retrieved
December 10, 2009,
www.uncommondescent.com/evolution/gettingover- our-love-for-darwin/
After years of trying to contend with Darwinists on this blog to pin them down with definitions of information and life and to get them to concede that they are defying several proven scientific laws when they defend Darwinism with absolutely no observed evidence, I am changing my tactics. My intention for the moment is to present various pieces of evidence that bulwark the idea of creation by a Creator God as the only way this Universe could exist in this form and that we could be alive.
Darwinists have their card tricks, just like people who look for water with a branched stick, known as water witching or dowsing. Magicians can make birds fly from hats and Darwinists can "discover" Tiktaalik the amazing transitional fossil that oops really isn't after all. No matter what they tell you, the sedimentary rock records are best explained by a worldwide flood and the ice age - mudrock age immediately thereafter. No matter what they tell you, Darwinists have no explanation for the beginning of the Universe or life or the source of information or design. They certainly cannot explain all sorts of symbiotic and non-essential behaviors and systems and structures found in nature that could not be a result of Darwinian evolution. The very makeup of DNA and the cell are impossible by Darwinist standards. The laws of thermodynamics are defied by Darwinism. Darwinism was once a working thesis that turned out to be a bad fit for reality but since it is now a column that upholds the humanist religion it must be defended by Darwinist zealots to the point of insanity. Because Humanism has become the state religion of the United States, Darwinism must be defended to the death by the high priests of the paradigm and all outbreaks of creationism or intelligent design must be stifled. It is not about science to them, it is about religious zealotry!
When one examines courses of study involving evolution, the idea of variation within kind by natural selection is always presented. However, this is a feature of baramin that allows for organisms to continue to survive and thrive as the ecosystem changes. The information to vary in kind is pre-existing within the genome, which means it was there from the beginning. No Darwinist can show you evolution happening, simply variation within kind. Then they will begin lying about the rock records and bring in their phylogeny phalsehoods that look good on the outside but at the micro level are proven to be false. Over the coming months I will be demonstrating this and also reminding readers of facilitated variation and genetic redundancy, two important features of reproduction and variation that support creation/ID and falsify Darwnism. If you do the research, you see that Darwin fails the smell test scientifically and philosophically as well.
If you believe in Darwinism, you believe we are random creatures in a random universe without meaning and without law. We have no compunction to do right or wrong for there is no basis for either. Selfishness is the only logical course of action. If there is no God and no standard of right and wrong, why not party hearty and grab for all the gusto you can get? Push weaker ones aside to get what you want and then take it all for yourself. Why not? Everyone is just going to die and that will be the end of that anyway. So why not live for today and screw the world?
Special creation, though, involves a Creator. The Creator has set a standard for living and has revealed Himself to his creatures. He has offered to share His knowledge and wisdom and has also extended an invitation to step beyond this physical realm into the supernatural upon death. What material things that have been created shall eventually run down to total entropy and without the actions of God heat death is the eventual end of the four dimensional Universe we experience. But God has created each of us with an undying self, whether you prefer to use the term spirit or soul, a self that will never cease to exist just as God will never cease to exist. You can choose to accept God's offer of salvation and mercy and fellowship and be upgraded to the supernatural life with Him in Heaven forever, or you will experience an undying destruction after death that is termed in the Bible as Hell.
Creationism works within the laws previously mentioned and follows the 2LOT as well. Only a Creator could have input the life and information and structures and systems in place to allow for life and could have built an entire Universe that follows logical laws and structures. Random chance would produce nothing but randomness. Entropy is produced from energy and not the other way around. Darwinists might as well go try to make a perpetual motion machine or discover the Philosopher's Stone or seek for the Holy Grail, their chances would be much better than they are to ever discover macroevolution in action. It isn't going to happen and it could not be an explanation for even one form of life on Earth, let alone millions!
When one examines courses of study involving evolution, the idea of variation within kind by natural selection is always presented. However, this is a feature of baramin that allows for organisms to continue to survive and thrive as the ecosystem changes. The information to vary in kind is pre-existing within the genome, which means it was there from the beginning. No Darwinist can show you evolution happening, simply variation within kind. Then they will begin lying about the rock records and bring in their phylogeny phalsehoods that look good on the outside but at the micro level are proven to be false. Over the coming months I will be demonstrating this and also reminding readers of facilitated variation and genetic redundancy, two important features of reproduction and variation that support creation/ID and falsify Darwnism. If you do the research, you see that Darwin fails the smell test scientifically and philosophically as well.
If you believe in Darwinism, you believe we are random creatures in a random universe without meaning and without law. We have no compunction to do right or wrong for there is no basis for either. Selfishness is the only logical course of action. If there is no God and no standard of right and wrong, why not party hearty and grab for all the gusto you can get? Push weaker ones aside to get what you want and then take it all for yourself. Why not? Everyone is just going to die and that will be the end of that anyway. So why not live for today and screw the world?
Special creation, though, involves a Creator. The Creator has set a standard for living and has revealed Himself to his creatures. He has offered to share His knowledge and wisdom and has also extended an invitation to step beyond this physical realm into the supernatural upon death. What material things that have been created shall eventually run down to total entropy and without the actions of God heat death is the eventual end of the four dimensional Universe we experience. But God has created each of us with an undying self, whether you prefer to use the term spirit or soul, a self that will never cease to exist just as God will never cease to exist. You can choose to accept God's offer of salvation and mercy and fellowship and be upgraded to the supernatural life with Him in Heaven forever, or you will experience an undying destruction after death that is termed in the Bible as Hell.
Creationism works within the laws previously mentioned and follows the 2LOT as well. Only a Creator could have input the life and information and structures and systems in place to allow for life and could have built an entire Universe that follows logical laws and structures. Random chance would produce nothing but randomness. Entropy is produced from energy and not the other way around. Darwinists might as well go try to make a perpetual motion machine or discover the Philosopher's Stone or seek for the Holy Grail, their chances would be much better than they are to ever discover macroevolution in action. It isn't going to happen and it could not be an explanation for even one form of life on Earth, let alone millions!
I am going to quit using the word "microevolution" as it is being consistently misused and misunderstood by the general public. This article (with bolding by me in some cases) explains why:
Variation, information and the created kind
Summary
All observed biological changes involve only conservation or decay of the underlying genetic information. Thus we do not observe any sort of evolution in the sense in which the word is generally understood. For reasons of logic, practicality and strategy, it is suggested that we:
Most popular literature on evolution more or less implies that since we see small changes going on today in successive generations of living things, we only have to extend this in time and we will see the types of changes which have caused single-cell-to-man evolution. Creationists are thus seen as drawing some sort of imaginary ‘Maginot line’, and saying in effect ‘this much variation we will allow but no more—call it microevolution or variation within kind.’ When a creationist says that, after all, mosquitoes are not seen turning into elephants or moths, this is regarded as a simplistic retreat. Such a criticism is not without some justification, because the neo-Darwinist can rightly say that he would not expect to see that sort of change in his lifetime either. The post-neo-Darwinist may say that our sample of geologic time is too small to be sure of seeing a ‘hopeful monster’ or any sort of significant saltational change.- Avoid the use of the term ‘microevolution’.
- Rethink our use of the whole concept of ‘variation within kind’.
- Avoid taxonomic definitions of the created kind in favour of one which is overtly axiomatic.
Another reason why the creationist position often appears as one of weakness is that we are perceived as admitting variation only because of being forced to do so by observation, then simply escaping the implications of variation by saying it does not go far enough. And we appear to redraw our ‘Maginot line’ depending on how much variation is demonstrated. It will be shown shortly, though, that this is a caricature of the creationist position, and that the limits to variation arise from basic informational considerations at the genetic level.
The created kinds
Observed variation does appear to have limits. It is tempting to use this fact to show that there are created kinds, and that variation is only within the limits of such kinds.However, the argument is circular and thus vulnerable. Since creationists by definition regard all variation as ‘within the limits of the created kind’ (see for example the statement of belief of the Creation Research Society of the USA), how can we then use observations to prove that variation is within the limits of the kind? To put it another way—of course we have never observed variation ‘across the kind’, since whatever two varieties descend from a common source, they are regarded as the same kind. It is no wonder that evolutionists are keen to press us for an exact definition of the created kind, since only then does our claim of ‘variation is only within the kind’ become non-tautologous and scientifically falsifiable.
Circular reasoning does not invalidate the concept of created kinds, however. In the same way, natural selection is also only capable of a circular definition (those who survive are the fittest, and the fittest are the ones who survive), but it is nevertheless a logical, easily observable concept. All we are saying is that arguments which are inherently circular cannot be invoked as independent proof of the kinds.
When I claim that such independent proof may not be possible by the very nature of things, this statement is in no way a ‘cop out’. For instance, let us say we happened upon the remnants of an island which had exploded, leaving behind the debris of rocks, trees, sand, etc. It may be impossible in principle to reconstruct the original positions of the pieces in relation to each other before the explosion. This does not, however, mean that it is not possible to deduce with a great degree of confidence that the current state of the debris is consistent with that sort of an explosion which was recorded for us by eyewitness testimony, rather than arising by some other mechanism.
In like manner, we can show that the observations of the living world are highly consistent with the biblically described concept of original created kinds, and inconsistent with the idea of evolution. This is best done by focusing on the underlying genetic/informational basis of all biological change. This is more realistic and more revealing than focusing on the degree or extent of morphological change.
The issue is qualitative, not quantitative. It is not that the train has had insufficient time to go far enough—it is heading in the wrong direction. The limits to variation—observed or unobserved—will come about inevitably because gene pools run out of ‘functionally efficient’ genetic information (or ‘teleonomic’ information). A full understanding of this eliminates the image of the desperately backpedalling creationist, redrawing his line of last resistance depending on what new observations are made on the appearance of new varieties.
It also defuses the whole issue of ‘micro’ and ‘macro’ evolution. I believe it is better for creationists to avoid these confusing and misleading terms altogether. The word ‘evolution’ generally conveys the meaning of the sort of change which will ultimately be able to convert a protozoon into a man or a reptile into a bird, and so on. I hope to show that in terms of that sort of meaning, we do not see any evolution at all. By saying ‘we accept micro but not macroevolution’ we risk reinforcing the perception that the issue is about the amount of change, which it is not. It is about the type of change.
This is not merely petty semantics, but of real psychological and tactical significance. Of course one can say that ‘microevolution’ occurs when this word is defined in a certain fashion, but the impact of the word, the meaning it conveys, is such as to make it unwise to persevere with this unnecessary concessional statement. Microevolution, that is, a change, no matter how small, which is unequivocally the right sort of change to ultimately cause real, informationally ‘uphill’ change, has never been observed.
In any case, leading biologists are themselves now coming to the conclusion that ‘macroevolution’ is not just ‘microevolution’ [using their terminology] extended over time. In November 1980 a conference of some of the world’s leading evolutionary biologists, billed as ‘historic’, was held at the Chicago Field Museum of Natural History on the topic of ‘macroevolution’. Reporting on the conference in the journal Science, Roger Lewin wrote:
The central question of the Chicago conference was whether the mechanisms underlying microevolution can be extrapolated to explain the phenomena of macroevolution. At the risk of doing violence to the positions of some of the people at the meeting, the answer can be given as a clear, No.1Francisco Ayala (Associate Professor of Genetics, University of California), was quoted as saying:
… but I am now convinced from what the paleontologists say that small changes do not accumulate.2The fact that this article reaches essentially the same conclusion in the following pages can thus hardly cause it to be regarded as radical. Nevertheless, the vast majority of even well-educated people still persist in ignorance of this. That is, they believe that ‘Big Change = Small Change x Millions of Years.’
The concept of information
The letters on this [printed] page—that is, the matter making up the ink and paper—all obey the laws of physics and chemistry, but these laws are not responsible for the information they carry. Information may depend on matter for its storage, transmission and retrieval, but is not a property of it. The ideas expressed in this article, for instance, originated in mind and were imposed on the matter. Living things also carry tremendous volumes of information on their biological molecules—again, this information is not a property of their chemistry, not a part of matter and the physical laws per se. It results from the order—from the way in which the letters of the cell’s genetic ‘alphabet’ are arranged. This order has to be imposed on these molecules from outside their own properties. Living things pass this information on from generation to generation. The base sequences of the DNA molecule effectively spell out a genetic ‘blue-print’ which determines the ultimate properties of the organism. In the final analysis, inherited biological variations are expressions of the variations in this information. Genes can be regarded as ‘sentences’ of hereditary information written in the DNA ‘language’.Imagine now the first population of living things on the evolutionist’s ‘primitive earth’. This so-called ‘simple cell’ would, of course, have a lot of genetic information, but vastly less than the information in only one of its present-day descendant gene pools, e.g., man. The evolutionist proposes that this ‘telegram’ has given rise to ‘encyclopedias’ of meaningful, useful genetic sentences. (See later for discussion of ‘meaning’ and ‘usefulness’ in a biological sense.) Thus he must account for the origin with time of these new and meaningful sentences. His only ultimate source for these is mutation.3
Going back to the analogy of the printed page, the information in a living creature’s genes is copied during reproduction, analogous to the way in which an automatic typewriter reproduces information over and over. A mutation is an accident, a mistake, a ‘typing error’. Although most such changes are acknowledged to be harmful or meaningless, evolutionists propose that occasionally one is useful in a particular environmental context and hence its possessor has a better chance of survival/reproduction. By looking now at the informational basis for other mechanisms of biological variation, it will be seen why these are not the source of new sentences and therefore why the evolutionist generally relies on mutation of one sort or another in his scheme of things.
1. Mendelian variation
This is the mechanism responsible for most of the new varieties which we see from breeding experiments and from reasonable inferences in nature. Sexual reproduction allows packets of information to be combined in many different ways, but will not produce any new packets or sentences. For example, when the many varieties of dog were bred from a ‘mongrel’ stock, this was achieved by selecting desired traits in successive generations, such that the genes or sentences for these traits became isolated into certain lines. Although some of these sentences may have been ‘hidden from view’ in the original stock, they were already present in that population. (We are disregarding mutation for the moment, since such new varieties may arise independently of any new mutations in the gene pool. Some dogs undoubtedly have mutant characteristics.)This sort of variation can only occur if there is a storehouse of such sentences available to choose from. Natural (or artificial) selection can explain the survival of the fittest but not the arrival of the fittest, which is the real question. These Mendelian variations tell us nothing about how the genetic information in the present stock arose. Hence, it is not the sort of change required to demonstrate ‘upward’ evolution—there has been no addition of new and useful ‘sentences’. And this is in spite of the fact that it is possible to observe many new varieties in this way—even new species. If you define a species as a freely interbreeding natural unit, it is easy to see how new species could arise without any ‘uphill’ change. That is, without the addition of any new information coding for any new functional complexity. For example, mutation could introduce a defect which served as a genetic barrier, or simple physical differences, such as the sizes of Great Dane and Chihuahua, could make interbreeding impossible in nature.
It is a little surprising to still see the occasional creationist literature clinging to the concept that no new species have ever been observed. Even if this were true, and there is some suggestion that it has actually been observed, there are instances of ‘clines’ in field observations which make it virtually certain that two now-isolated (reproductively) species have arisen from the same ancestral gene pool. Yet the very same creationists who seem reluctant to make that sort of admission would be quite happy to agree with the rest of us that the various species within what may be regarded as the ‘dog’ kind, including perhaps wolves, foxes, jackals, coyotes and the domestic dog, have arisen from a single ancestral kind. So why may this no longer be permitted to be happening under present-day observations? It is not only biblically and scientifically unnecessary, but it sets up a ‘straw man’ in the sense that any definite observation of a new species arising is used as a further lever with which to criticize creationists.
What we see in the process of artificial selection or breeding giving rise to new varieties, is a thinning-out of the information in the parent stock, a reduction in the genetic potential for further variation. If you try and breed a Chihuahua from a Great Dane population or vice versa, you will find that your population lacks the necessary ‘sentences’. This is because, as each variety was selected out, the genes it carried were not representative of the entire gene pool.
What appeared to be a dramatic example of change with the appearance of apparently new traits thus turns out, when its genetic basis is understood, to be an overall downward movement in informational terms. The number of sentences carried by each subgroup is reduced thus making it less likely to survive future environmental changes. Extrapolating that sort of process forward in time does not lead to upwards evolution, but ultimately to extinction with the appearance of evermore-informationally-depleted populations.
2. Polyploidy
Again, no sentences appear which did not previously exist. This is the multiplication (‘photocopying’) of information already present.3. Hybridizatlon
Again, no new sentences. This is the mingling of two sets of information already present.4. Mutation
Since mutations are basically accidents, it is not surprising that they are observed to be largely harmful, lethal or meaningless to the function or survival of an organism. Random changes in a highly ordered code introduce ‘noise’ and chaos, not meaning, function and complexity, which tend to be lost. However, it is conceivable that in a complex world, occasionally a ‘destructive’ change will have a limited usefulness. For example, if we knock out a sentence such that there is a decrease in leg length in sheep (and there is such a mutation), this is useful to stop them jumping over the farmer’s fence. A beetle on a lonely, wind-swept island may have a mutation which causes it to lose or corrupt the information coding for wing manufacture; hence its wingless successors will not be so easily blown out to sea and will thus have a selective advantage. Eyeless fish in caves, some cases of antibiotic resistance—the handful of cases of mutations which are quite ‘beneficial’—do not involve the sort of increase in functional complexity which evolutionary theory demands. Nor would one expect this to be possible from a random change.At this point some will argue that the terms ‘useful’, ‘meaningful’, ‘functional’, etc. are misused. They claim that if some change gives survival value then by definition it has biological ‘meaning’ and usefulness. But this assumes that living systems do nothing but survive—when in fact they and their subsystems carry out projects and have specific functions. That is, they carry teleonomic information. This is one of the essential differences between living objects and non-living ones (apart from machines). These projects do not always give rise to survival/reproductive advantages—in fact, they may have very little to do with survival, but are carried out very efficiently. The Darwinian assumption is always made, of course, that at some time in the organism’s evolutionary history, the project had survival/reproductive value. (For example, the archer-fish with its highly-skilled ‘hobby’ of shooting down bugs which it does not require for survival at the present time.) However, since these are nontestable assumptions, it is legitimate to talk about genetic information in a teleonomic sense, in isolation from any possible survival value.
The gene pools of today carry vast quantities of information coding for the performance of projects and functions which do not exist in the theoretical ‘primeval cell’. Hence, in order to support protozoon-to-man evolution, one must be able to point to instances where mutation has added a new ‘sentence’ or gene coding for a new project or function. This is so regardless of one’s assumptions on the survival value of any project or function.
We do not know of a single mutation giving such an increase in functional complexity. Probabilistic considerations would seem to preclude this in any case, or at least make it an exceedingly rare event, far too rare to salvage evolution even over the assumed multibillion year time span.
To illustrate further—the molecule haemoglobin in man carries out its project of transporting and delivering oxygen in red cells in a functionally efficient manner. A gene or ‘sentence’ exists which codes for the production of haemoglobin. There is a known mutation (actually three separate ones, giving the same result) in which only one letter in the sentence has been accidentally replaced by another. If you inherit this change from both parents, you will be seriously ill with a disease called sickle cell anaemia and will not survive for very long. Yet evolutionists frequently use this as an example of a ‘beneficial mutation’. This is because if you inherit it from only one parent, your red cells will be affected, but not seriously enough to affect your survival—just enough to prevent the malaria parasite from using them as an effective host. Hence, you will be more immune to malaria and better able to survive in malaria-infested areas. This shows us how a functionally efficient haemoglobin molecule became a functionally crippled haemoglobin molecule. The mutation-caused gene for this disease is maintained at high levels in malaria-endemic regions by this incidental phenomenon of heterozygote superiority. Its damaging effect in a proportion of offspring is balanced by the protection it gives against malaria. It is decidedly not an ‘upward’ change. We have not seen a new, efficient oxygen transport mechanism or its beginnings evolve. We have not seen the haemoglobin transport mechanism improved.
One more loose but possibly useful analogy. Let us say an undercover agent is engaged in sending a daily reassuring telegram from enemy territory. The text says ‘the enemy is not attacking today’. One day an accident occurs in transmission and the word ‘not’ is lost. This is very likely going to be a harmful change, perhaps even triggering a nuclear war by mistake. But perhaps, in a freak situation, it could turn out to be useful (for example, by testing the fail-safe mechanisms involved). But this does not mean that it is the sort of change required to begin to convert the telegram into an encyclopedia.
The very small number of ‘beneficial’ mutations actually observed are simply the wrong kind of change for evolution—we do not see the addition of new sentences which carry meaning and information. Again surprisingly, one often reads creationist works which insist that there is no such thing as a beneficial mutation. If benefit is defined purely in survival terms, then we would not expect this to be true in all instances, and in fact it is not—that is, there are indeed ‘beneficial’ mutations in that sense only.
Information depends on order, and since all of our observations and our understanding of entropy tells us that in a natural, spontaneous, unguided and unprogrammed process order will decrease, the same will be true of information. The physicist and communications engineer should not be surprised at the realisation that biological processes involve no increases in useful or functional (teleonomic) information and complexity. In fact, the net result of any biological process involving transmission of information (i.e., all hereditary variation) is conservation or loss of that genetic information.
This points back directly to the creation of the information, supernaturally, in the beginning. It is completely in harmony with the biblical concept of a world made ‘very good’ as a balanced, functioning whole, with decay only subsequent to the Fall. This is the reason why there are inevitable limits to variation, why the creationist does not have to worry about how many new ‘species’ the future may bring—because there is a limit to the amount of functionally efficient genetic information present, and natural processes such as mutation cannot add to this original storehouse.
Notice that since organisms were created to migrate out from a central point at least once and fill empty ecological niches, as well as having to cope with a decaying and changing environment, they would require considerable variation potential. Without this built-in genetic flexibility, most populations would not be present today. Hence the concept of biological change is in a sense predicted by the biblical model, not something forced upon it only because such change has occurred.
The created kind
The Scriptures imply that this originally created information was not in the form of one ‘super species’ from which all of today’s populations have split off by this ‘thinning out’ process, but was created as a number of distinct gene pools. Each group of sexually reproducing organisms had at least two members. Thus,- Each original group began with a built-in amount of genetic information which is the raw material for virtually all subsequent useful variation.
- Each original group was presumably genetically and reproductively isolated from other such groups, yet was able to interbreed within its own group. Hence the original kinds would truly have earned the modern biological definition of ‘species’.4 We saw in our dog example that such ‘species’ can split into two or more distinct subgroups which can then diverge (without adding anything new) and can end up with the characteristics of ‘species’ themselves—that is, reproductively isolated from each other but freely interbreeding among themselves. The more variability in the original gene pool, the more easily can such new groups arise. However, each ‘splitting’ reduces the potential for further change and hence even this is limited. All the descendants of such an original kind which was once a species, may then end up being classified together in a much higher taxonomic category—e.g., family.
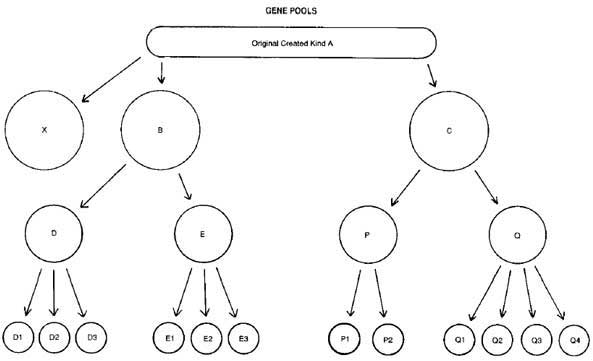 Figure 1. The ‘splitting off’ of daughter populations from an original created kind. Click here for larger image. |
Hence, D1, D2, D3, E1, E2, E3, P1, P2, Q1, Q2, Q3 and Q4 are all different species, reproductively isolated. But all the functionally efficient genetic information they contain was present in A. (They presumably carry some mutational defects as well).
Let us assume that the original kind A has become extinct, and also the populations X, B, C, D, E, P and Q. (But not D1, D2, etc.) If X carried some of the original information in A, which is not represented in B or C, then that information is lost forever. Hence, in spite of the fact that there are many ‘new species’ which were not originally present, we would have witnessed conservation of most of the information, loss of some, and nothing new added apart from mutations (harmful defects or just meaningless ‘noise’ in the genetic information). All of which is the wrong sort of informational change if one is trying to demonstrate protozoon-to-man evolution.
Classifications above species are more or less arbitrary groupings of convenience, based generally on similarities and differences of structure. It is conceivable that today, D1, D2 and D3 could be classified as species belonging to one genus, and E1, E2 and E3 as species in another genus, for example. It could also be that the groups B and C were sufficiently different such that their descendants would today be in different families. We begin to see some of the problems facing a creationist who tries to delineate today’s representatives of the created kinds.
Creatures may be classified in the same family, for example, on the basis of similarities due to common design while in fact they belong to two totally different created kinds. This should sound a note of caution against using morphology alone, as well as pointing out the potential folly of saying ‘in this case, the baramin is the family; in this case, it is the genus, etc.’ (Baramin is an accepted creationist term for ‘created kind’.)
There is no easy solution as yet to the problem of establishing each of these genetic relationships—in fact, we will probably never be able to know them all with certainty. Interbreeding, in vitro fertilization experiments, etc. may suggest membership of the same baramin but lack of such genetic compatibility does not prove that two groups are not in the same kind. (See earlier discussion—genetic barriers could arise via mutational deterioration.) However, newer insights, enabling us to make direct comparisons between species via DNA sequencing, open up an entirely new research horizon. (Although the question of where the funding for such extensive research will come from in an evolution-dominated society remains enigmatic.)
What then do we say to an evolutionist who understandably presses us for a definition of a created kind or identification of same today? I suggest the following for consideration:
Groups of living organisms belong in the same created kind if they have descended from the same ancestral gene pool.To talk of ‘fixity of kinds’ in relation to any present-day variants thus also becomes redundant—no new kinds can appear by definition.
Besides being a simple and obvious definition, it is axiomatic. Thus it is as unashamedly circular as a rolled-up armadillo and just as impregnable, deflecting attention, quite properly, to the real issue of genetic change.
The question is not—what is a baramin, is it a species, a family or a genus? Rather, the question is—which of today’s populations are related to each other by this form of common descent, and are thus of the same created kind? Notice that this is vastly removed from the evolutionist’s notion of common descent. As the creationist looks back in time along a line of descent, he sees an expansion of the gene pool. As the evolutionist does likewise, he sees a contraction.
As with all taxonomic questions, common sense will probably continue to play the greatest part. The Scriptures, the fossil record and common sense unite to prevent creationists doing too much ‘lumping together’ as we go back in time. For instance, it is conceivable (though not necessarily so) that crocodiles and alligators both descended from the same ancestral gene pool which contained all their functionally efficient genes, but not really conceivable that crocodiles, alligators and ostriches had a common ancestral pool which carried the genes for all three!
Related articles
Footnotes
- Lewin, R., Evolutionary theory under fire, Science 210 (4472): 883–887, 1980. Return to text.
- Lewin, Ref. 1, p. 884. Return to text.
- Transposons or ‘jumping genes’, which involve the transfer of genetic information from one stretch of DNA to another (in the same organism, or even altogether different ones, transferred by viruses for example) are not touched upon in this discussion, as work on these ‘new hopes’ for evolution is not at sufficiently mature stage. It should, however, be noted that these processes would appear to be (like mutations) random and undirected processes, in this case involving transfers of existing information. Return to text.
- Of course, the original ‘kind’ may have been represented by two or more separate populations able to interbreed potentially, but geographically isolated. Thus, being reproductively ‘isolated’ would cause them, by the ultra-strict application of the definition, to be more than one ‘species’ in the ‘kind’. Some creationists would call these separate ‘morphotypes’ within each baramin as originally created.
The article above has convinced me to give up the word "microevolution" and stick to "variation within kind", which is both more specific and less confusing. Baramin, by the way, comes from Bara (created) and Min (kind) so a Min is a kind but all Min were created at the beginning and the word from the Bible is Baramin, thus, Baraminology!
Baraminology is a scientific way to sort distinct kinds of animals and their subgroups. Linnaeus was attempting to do this with his classification system, which has been taken by Darwinists and twisted and sometimes laughably broken. There really is no word (family, genus, species) that works as well or is as specific as a baramin. All wolf-dog types are of one baramin, for instance.
"For instance, it is conceivable (though not necessarily so) that crocodiles and alligators both descended from the same ancestral gene pool which contained all their functionally efficient genes, but not really conceivable that crocodiles, alligators and ostriches had a common ancestral pool which carried the genes for all three!"
Two important quotes from the telling article from 2009 listed below:
(Concerning the British public's tendency to believe in Intelligent Design) "Or you may do so because you genuinely do not know of the evidence for evolution, have never had it explained to you, or because you just don’t understand it."
"It" being the hypothesis of evolution by mutations in combination with natural selection over long periods of time. Of course the evidence for Darwinism is all a matter of just-so stories, it is not even possible for natural selection to operate on organisms that do not yet exist and there is no means by which organisms can form from matter naturally. Darwin has no explanation for life or information so it is dead in the water from the beginning. It also runs in the wrong direction, as the article above illustrates.
“This is a good thing, because as evolution is arguably the most important concept underlying the life sciences,” he said, “providing children with an understanding of it [evolution] at the earliest possible age will surely help lay the foundations for a surer scientific understanding later on.”
Balderdash! Humbug! Tommyrot! Lies! Foolishness! Darwinism has been nothing but a hindrance to operational science. How many tonsils were removed when they might better have been kept?. How much medical knowledge was held back because researchers believed in the idea of vestigal organs? We now know of no vestigal structures in humans at all. Darwinism was and is a drag on science.
Today we have millions of dollars wasted scanning the skies for a message from ET and think of the wasted salaries of every single member of the NCSE, which is nothing more than a Darwinist censorship organization!
Researchers have to throw in an homage to Darwin somewhere early in their publications before then going into the nuts and bolts of research than do not utilize Darwin at all. If we took Darwin out of science, the research in real science would continue unabated. Only the people trying in vain to make life come from non-life, make apes appear to be people or furiously writing dumbed-down explanations for textbooks would be out of a job!
From the November/December Creation Matters Magazine:
Darwin Marketed to Kids (Great Britain)
There’s a move on to get Darwin’s ideas taught to kids. Britain is giving a “birthday present to Darwin,”
wrote Andrew Copson for The Guardian,1 in the form of a national curriculum for primary schools
that will mention evolution for the first time — and prohibit teaching of creationism or intelligent design
in science lessons.
The addition of evolution to elementary school curriculum was in response to a letter promoted by the
British Humanist Association and signed by “scientists and experts.”Copson was obviously delighted
with what he perceived as a long-overdue smackdown against intelligent design — a belief espoused
by the majority of his fellow Britons: Those who care about public reason are routinely shocked by opinion
polls and surveys showing high levels of credence given to the idea of intelligent design. The most
recent poll purported to demonstrate that a majority of Britons think that it should be taught alongside evolution
in schools.
To solve this problem, we have to know what causes it and there are two reasons why you might prefer
the idea of intelligent design to that of evolution. You may do so because your prior ideological convictions,
mostly to do with religious belief, simply don’t allow you to accept the evidence that is presented to you.
Or you may do so because you genuinely do not know of the evidence for evolution, have never had it
explained to you, or because you just don’t understand it. In a society as decreasingly religious as England,
it is impossible to believe that most of the people who do not accept evolution are motivated by ideology rather than ignorance. This means that the best way to solve the problem is through better education and that is
what makes the inclusion of evolution in the science curriculum as early as possible so important.
Repeatedly in his article, Copson characterized the Darwin doubters as ignorant, but refused to acknowledge any ideological bias on his part. He also made it seem as if all evidence is for evolution and against religion — despite numerous evidential claims by intelligent design against evolution. He spoke favorably of a slate of
new children’s books on evolution such as What Mr. Darwin Saw, How Whales Walked into the Sea, and Mammals Who Morph.
“This is a good thing, because as evolution is arguably the most important concept underlying the life sciences,” he said, “providing children with an understanding of it [evolution] at the earliest possible age will surely help
lay the foundations for a surer scientific understanding later on.”
A new company is making Darwin toys for tots. Charlie’s Playhouse2 offers “Evolution for kids” in the
form of apparel, games, cards, and a giant evolution timeline kids can hop and skip on. Of special interest
is their 30-second commercial, Why are we making evolution toys? The answer: nobody else is. The
commercial laments the thousands of toys about physics, biology, and chemistry — even all those popular dinosaur toys — that don’t mention evolution. “But we do!” a cartoony Darwin exclaims, dancing
proudly at the end of the video.
William Dembski, the double-PhD scholar of the intelligent design movement, has had enough with all this.
His latest article on Uncommon Descent3 is called “Getting over our love for Darwin.” In it he quotes
Malcolm Muggeridge, who wrote, “I myself am convinced that the theory of evolution, especially the extent to which it’s been applied, will be one of the great jokes in the history books in the future. Posterity will marvel that so very flimsy and dubious an hypothesis could be accepted with the incredible credulity that it has.”
Exactly
1. Copson, A. (2009, November 9). A birthday present
for Darwin. Guardian. Retrieved December
10, 2009, from
www.guardian.co.uk/commentisfree/belief/2009 /nov/09/evolution-primary-schools-science
2. Charlie’s Playhouse. 2009. Why are we making
evolution toys? Retrieved December 20, 2009,
from http://www.charliesplayhouse.com/whyevolution- toys.php
3. Dembski, W.A. (2009, November 3). Getting over
our love for Darwin. Uncommon Descent. Retrieved
December 10, 2009,
www.uncommondescent.com/evolution/gettingover- our-love-for-darwin/