The Earth is how old? Says who? Oh.
Before I post the follow up to the "take my primordial soup, please" post, I wanted to share this information that came from the CMI site. Creation.com is the site where one of the prime contributors is Jonathan Sarfati, the guy Richard Dawkins and other Darwinists are afraid to debate (and for good reason) and several other top flight scientists who do NOT believe in Chance, the Evolution Fairy.
Old-earth or young-earth belief
Which belief is the recent aberration?
CMI takes the stand that the creation is thousands of years old, based on straightforward acceptance of the chronology in Genesis.Churchian critics accept the billions of years touted today and claim that the ‘young-earth’ view is a recent invention of Protestant ‘fundamentalist’ churches. They claim that various Church Fathers or other ancient authorities wrote things that suggested they did not hold a ‘young-earth’ view.
However, such claims about the Church Fathers and Reformers have been shown to be wrong; for instance, in the rebuttal to the long-ager Hugh Ross, by Mark Van Bebber and Paul Taylor.1 It’s vital to analyse what the scholars of the past actually said, instead of what people claim they said2—see Did early Church leaders and reformers believe the literal creation account given in Genesis? Sometimes a non-specific statement from them about Creation is misinterpreted as claiming that they didn't have a position on the days and timeframe. This is obviously the fallacy of arguing from silence.
It’s worse when clear statements about the Creation days and timeframe are ignored in favor of non-specific ones. The correct practice is to interpret the non-specific passages by the specific ones. Note that, by the same methodology, hundreds of years in the future, someone could find articles by every CMI writer that are not specific on days or the timeframe and, by ignoring their clear statements elsewhere, claim that we don’t have any position on these issues!
Even Augustine cannot remotely be used in support of old-earth beliefs. Even though he allegorized the Days of Creation (and lots of other passages—he was no Hebrew scholar), he tried to compress the days into an instant, which is diametrically opposite to what long-agers claim!
Furthermore, when ancient chronologies are researched, we find that many cultures, not just those based directly on the Bible, attest to an age for the creation of thousands of years. It seems that no serious scholar believed in the old-earth fashion of today. It is very much a modernist invention.
The following comes from Young’s Analytical Concordance to the Holy Bible, first published in 1879. Under ‘Creation’, Young listed dates of Creation compiled by a Dr William Hales,3 who was an expert in chronology. None of them give a date for Creation of more than 9,000 years ago. Note that dates of Creation dates from various non-Christian/non-Jewish sources (India, Egypt, China, pre-Christian Greece, Babylonia, etc.) all testify to an age of thousands of years.4 Furthermore, both Catholic and Protestant scholars agreed on this issue. It seems that no serious chronologist believed in an old earth.
Many of the scholars listed in our table (below) made the chronology of the world a life-long study, involving meticulous cross-matching of ancient records (many of which are not available to us today). These scholars did not obtain these dates by throwing dice! It is only the arrogance of modern man that dismisses this tradition of careful scholarship.
We must underline again the importance of this issue. The old-earth advocates accept the recently fashionable claim that the earth is billions of years old. This ‘age’ is based on radioactive dating. The same ‘dating’ puts the age of fossilized dead animals at up to hundreds of millions of years before people appeared on the scene. These fossils show evidence of violence and suffering—cancer and arthritis can be seen in the fossils, for example (see Genesis: the Curse). So in the old-earth schemes, this death and suffering was going on when God created Adam and Eve, and pronounced everything as ‘very good’ (Genesis 1:31). Hardly!
The Bible clearly teaches that the bad things came into being when Adam and Eve rejected the rule of the Creator over them, and the Curse of death came into effect (Genesis 3:17–19, Romans 8:20–22, etc.). And because of this true history of the world, Jesus, the ‘last Adam’, came into the world to die for the sinful descendants of Adam (Romans 5:12 ff., 1 Cor. 15:21–22, 45).
In short, all attempts to marry the Bible with the old-earth view actually undermine the Gospel.
Date of Creation
(from Young’s Analytical Concordance of the Holy Bible, 1879 8th Edition, 1939—entry under ‘Creation’)‘Dr Hales, in his work entitled, “A New Analysis of Chronology and Geography, History and Prophecy,”? (vol. 1, p. 210 [published in 1830]), remarks: “In every system of chronology, sacred and profane, the two grand eras—of the Creation of the World, and of the Nativity of Christ—have been usually adopted as standards, by reference to which all subordinate epochs, eras and periods have been adjusted.”? He gives a list of 120 dates, commencing B.C. 6984, and terminating BC 3616, to which this event has been assigned by different authorities, and he admits that it might be swelled to 300. He places it at BC 5411. The date commonly adopted is BC 4004; being that of Ussher, Spanheim, Calmet, Blair, etc., and the one used in the English Bible [KJV].’
‘The following are some of the principal variations:’ (notations regarding dates of compilation or publication, extra information on sources, etc., have been added by CMI).
| Source of Creation Date | Authority | Date BC |
|---|---|---|
| Alfonso X (Spain, 1200s) | Muller | 6984 |
| Alfonso X (Spain, 1200s) | Strauchius, Gyles (1632–1682)5 | 6484 |
| India | Gentil, French astronomer c.1760 | 6204 |
| India | Arab records | 6174 |
| Babylonia | Bailly6 (French astronomer, 1736–1793) | 6158 |
| China | Bailly | 6157 |
| Diogenes Laertius (Greece 3rd Cent.) | Playfair | 6138 |
| Egypt | Bailly | 6081 |
| Septuagint (LXX)7 | Albufaragi | 5586 |
| Josephus (1st Century Jew) | Playfair | 5555 |
| Septuagint, Alexandrine | Scaliger, Joseph (1540–1609)8 | 5508 |
| Persia | Bailly | 5507 |
| Chronicle of Axum, Abyssinian | Bruce (1700s) | 5500 |
| Josephus | Jackson | 5481 |
| Jackson | 5426 | |
| Hales | 5411 | |
| Josephus (1st Century Jew) | Hales | 5402 |
| India | Megasthenes,9 Greek historian (c. 340–282 bc) | 5369 |
| Talmudists | Petrus Alliacens | 5344 |
| Septuagint, Vatican | 5270 | |
| Bede (673–735) | Strauchius | 5199 |
| Josephus (1st Century Jew) | Univ. Hist. | 4698 |
| Samaritan computation | Scaliger | 4427 |
| Samaritan text | Univ. Hist. | 4305 |
| Hebrew (Masoretic) text | 4161 | |
| Playfair and Walker | 4008 | |
| Ussher, Spanheim, Calmet, Blair, etc. | 4004 | |
| Kepler10 (1571–1630) | Playfair | 3993 |
| Petavius (France, 1583–1652) | 3984 | |
| Melanchthon (1500s) | Playfair | 3964 |
| Luther (1500s) | 3961 | |
| Lightfoot | 3960 | |
| Cornelius a Lapide | Univ. Hist. | 3951 |
| Scaliger, Isaacson | 3950 | |
| Strauchius | 3949 | |
| Vulgar Jewish computation | Strauchius | 3760 |
| Rabbi Lipman (1579–1654) | Univ. Hist. | 3616 |
‘Basil the Great’ on GenesisBasil was Bishop of Caesarea Mazaca, Cappadocia, from AD 370–379. He argued strongly against various heresies of the church of that day and was one of the great defenders of the doctrine of the Trinity. He also established charitable institutions, hospitals and schools, and organized famine relief. Basil was greatly admired and respected, and was called ‘Basil the Great’.Basil is one of the ‘Church Fathers’ that some have claimed did not believe in a historical Genesis. However, in a series of sermons on the Days of Creation, the Hexaëmeron (‘Six Days’), Basil showed that he believed:
NoteAdapted from David Watson, An early View of Genesis One, Creation Research Society Quarterly 27(4):138–139, 1991. Based on: Stanislaus Giet (translator), Homelies sur L'Hexaëmeron by St. Basil, originally written in AD 370. In the series Sources Chrétiennes, Les Editions du Gerf, Paris, 1949. |
References and Notes
- Creation and Time—a Report on the Progressive Creationist Book by Hugh Ross, Eden Productions, Mesa, Arizona, USA, 1994. Return to text.
- See Anon., Genesis means what it says—Basil (AD 329–379), Creation 16(4):23, 1994. Did early church leaders and Reformers believe the literal Creation account given in Genesis?, 9 July 2001. Return to text.
- published in 1830. Return to text.
- That is not to say that there were not beliefs in long ages, for example, with Greek and Indian mythological religions, as distinct from properly studied chronologies. Return to text.
- Brevarium Chronologicum Book IV, 3rd edition, 1699 in English. Return to text.
- Bailly, Jean Sylvain. Return to text.
- Based on the Greek translation of the Old Testament originally published in Egypt c. 250 BC. Return to text.
- French classical scholar. Return to text.
- A Greek historian from Iona, Megasthenes was Ambassador to India for King Seleucus I. He published Indika in four books. Return to text.
- Kepler was a Lutheran and A great scientist who discovered the laws of planetary motion. Return to text.
If you are unfamiliar with Creation.com, they are a science site with scientists engaged in research - operational science - the kind based upon evidence? Like this below, with findings that should not surprise those who have read the articles on facilitated variation and genetic redundancies that I have posted in the past year or so...Organisms are pre-configured to make certain choices and take on certain characteristics with the help of pre-existing switches within the organism as coded by DNA:
Karyotypic and allelic diversity within the canid baramin (Canidae)
Previous studies suggest that all dog-like creatures (canids, family Canidae) belong to a single created kind. As unclean animals, all modern canids are descendants of two canids preserved on the Ark during the Flood. This pair of canids would have carried a limited amount of genetic diversity. They would be expected to have had a fairly uniform arrangement of chromosomes (low karyotypic diversity) and up to four different versions of any particular gene (allelic diversity). Today there is considerably more karyotypic and allelic diversity within the canids. The patterns imply that more than random mutation and natural selection are involved; instead, certain genetic components appear designed to change and numerous designed mechanisms may be involved in driving many of these changes. This suggests that God, in His infinite wisdom, designed animals to be able to undergo certain genetic mutations which would enable them to adapt to a wide range of environmental challenges while minimizing risk.

Table 1. List of canid species and their normal diploid (2n) number which were included in a phylogenomic analysis by Graphodatsky et al.7
Previous baraminic studies have identified all canids (family Canidae) as belonging to a single baramin.3 Since they are unclean animals, all living canids would have descended from a single breeding pair preserved on the Ark about 4,500 years ago.4,5 This historical information is important because it suggests there was a limited amount of diversity present in canids at that time. Today, this family is represented by 34 species that are widely distributed around the world.6 There are considerable data available on the karyotypic and allelic diversity in protein coding genes for several of these species. A brief overview of the data is presented here.
By examining intrabaraminic genetic diversity, we should be able to discover a clearer picture regarding the role of mutations in the development of the diversity found in animals today.
Karyotype
The family Canidae exhibits the most highly rearranged karyotypes* of any family within the order Carnivora. Normal diploid numbers vary from 34 for the red fox (Vulpes vulpes) to 78 for the domestic dog (Canis familiaris) and dhole (aka Asiatic Wild Dog; Cuon alpinus) (table 1). The Arctic fox (Alopex lagopus) is polymorphic for a centric fusion; diploid numbers of 49 and 48 are found in individuals carrying one or two copies respectively of this fusion. Phylogenomic analysis suggests that 82 may have been the ancestral karyotype. Within the 10 species that have been studied in detail it appears that approximately 80 rearrangements have occurred. This includes numerous fusions, both centric and tandem, fissions, pericentric inversions and/or centromere transpositions.7 Several paracentric inversions, and even whole arm (telomere to centromere) inversions, have been implicated based on the differences in loci order among species (figure 1).8,9
Figure 1. Diagrams depicting some of the chromosomal rearrangements reported within the canid baramin. Such rearrangements often result in the loss of relatively small portions of DNA. Fusions (top row) involve combining two distinct chromosomes to form one; to become stable, one centromere must then be silenced. Inversions (bottom row) involve reorienting a portion of DNA within an existing chromosome. There also is evidence that the amount of heterochromatin can be adjusted. These types of rearrangements are too complex to be the result of ‘purely chance events’. While rearrangements do involve some risk, they probably also have purpose, such as adaptation in a fallen world.
Instead, the presences of multiple designed mechanisms enabling translocations to occur while maintaining viability of the animal suggests that such rearrangements are likely helpful for adaptation in the present fallen world.
Major histocompatibility complex genes
The major histocompatibility complex (MHC) consists of a number of genes involved in immune function and which are known for high allelic diversity. Several dog leukocyte antigen (DLA) genes have been evaluated for polymorphisms. As of 2006, there were 90 alleles recognized for DLA-DRB1, 22 for DLA-DQA1 and 54 for DLA-DQB1, with more expected to be discovered.17 High levels of polymorphism are generally considered a sign of a healthy population, although some dog breeds and wild mammals have low MHC diversity with no apparent ill effects. The DLA genes are on dog chromosome (CFA) 12.18 Some DLA haplotypes are associated with various canine autoimmune diseases such as primary immune mediated hemolytic anemia, polyarthritis, hypothyroidism and diabetes.19 However, it is important to recognize that these haplotypes do not cause disease directly; instead, they may be risk factors that affect the likelihood of disease development. As suggested previously, there is risk in maintaining sufficient variability to adapt in the present fallen world.Dopamine receptor D4 gene
There are two portions of the dopamine receptor D4 (DRD4) gene that are variable in dogs. The first is in exon 1 where the two known alleles differ by a 24-base pair (bp) indel.20 Interestingly, humans also are polymorphic in this region with a 12-bp duplication and a 13-bp deletion having been identified.21 The latter is particularly intriguing as it is found in 2% of the human population and is not associated with any known disease; yet the frameshift is predicted to result in a truncated, non-functional protein.22
Figure 2. A representation of the variable number tandem repeat (VNTR) patterns in exon 3 of the dopamine receptor D4 (DRD4) gene for seven dog alleles (after Hejjas et al.23). The nonrandom pattern of mutation suggests designed mechanisms are involved in this mutation. The variability in this region appears to have some influence on personality and behaviour.
This bias of indels, particularly VNTRs, in base pairs that are multiples of three does not appear to be explicable by natural selection. If essentially random, approximately one-third of indels should be multiples of three unless a frameshift, which often results in a premature stop codon and a nonfunctional protein, is lethal or significantly detrimental. It does not appear that frameshifts in DRD4 would be subject to such selection pressure, since a frameshift mutation is carried by a number of normal humans and knock-out mice.20,22 Furthermore, variability in this gene appears to contribute to variability in personality. The number of alleles in canids (greater than eight, as the raccoon dog has a separate allele identified25) is greater than the maximum of four alleles expected in the pair of canids on the Ark. Humans also carry more alleles than can be attributed to Adam and Eve. This suggests that this gene was designed to vary in a rather unusual way to enhance variability in personality and perhaps other traits as well.
Olfactory genes
Olfactory (smell) receptor (OR) genes are seven transmembrane receptors. While 1,094 OR genes have been identified in the dog,26 the canine repertoire of odorant molecules is significantly greater than this. This appears to be from a complex combinatorial code. Odorant molecules can bind 20 or more ORs depending on their concentration. ORs can bind more than one odorant molecule. Through interpretation of the complex signalling patterns, dogs are able to detect an incredibly wide array of individual odorants and a large number of mixtures.27In one study, 16 OR genes were examined in 95 dogs from 20 different breeds. All genes were polymorphic ranging from two to 11 alleles per gene. There was an average of one change per 920 sequenced nucleotides, which is much higher than most coding sequences and a random sampling of non-coding sequences. Of the 98 single nucleotide polymorphisms (SNPs) identified, 55 resulted in an amino acid change and 30 of these involved changes to a different amino acid group. These changes were found throughout the protein (figure 3), mostly in variable or highly variable regions within OR genes. However, two come from highly conserved regions, one in transmembrane (TM) 3 and the other in TM7.28
There is a tremendous amount of redundancy in OR [olfactory receptor] genes which may have been designed to allow for future specialization.
It may be premature to assume there is no purpose in mutation or pseudogenization within OR genes.29 There is a tremendous amount of redundancy in OR genes which may have been designed to allow for future specialization. For example, a study involving Drosophila sechellia, a highly specialized vinegar fly that feeds solely on fruit from Morinda citrifolia, a shrub which strongly repels related species of flies, suggests that pseudogenization of ORs and gustatory (taste) receptors has occurred nearly 10 times faster than in the closely related species D. simulans. For those genes which remained intact, D. sechellia appears to have fixed non-synonymous substitutions at a consistently higher rate than synonymous substitutions compared to the same genes in D. simulans.30 Therefore, the ability of OR genes to be modified or pseudogenized may be an important design element introduced by the Creator.
Conclusion
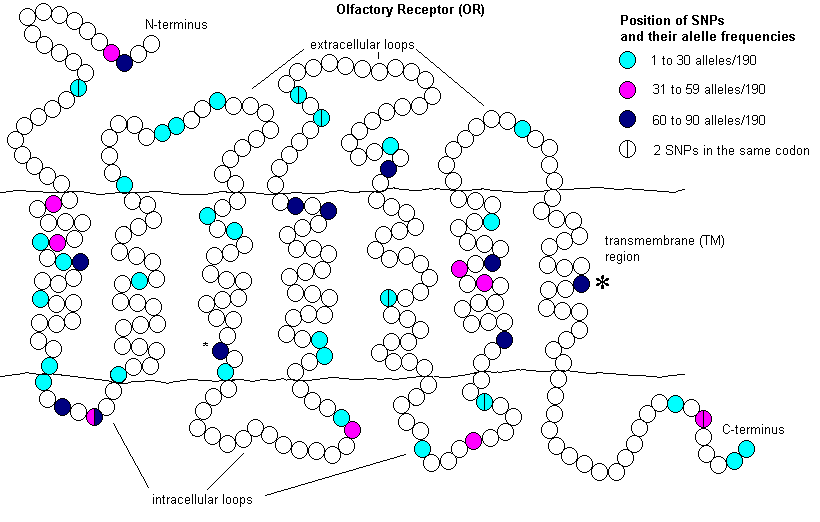
Figure 3. Two-dimensional diagram of an olfactory receptor (OR) indicating positions of 55 non-synonymous single nucleotide polymorphisms (SNPs) and their allele frequencies in dogs, as identified by Tacher et al.28 ‘*’ indicates the SNPs found in highly conserved regions of the OR genes. There are 1,094 OR genes that have been identified in dogs.
The various genes examined here appear to handle mutations very well. In fact, it is generally believed that the high allelic diversity in the MHC genes is important for a healthy population. The redundancy in ORs and the pattern of mutation and pseudogenization in these genes suggests that these genes were designed to vary so that animals can adapt to different environments. Finally, the striking non-random pattern of VNTR mutations, all in lengths divisible by three, when there is no known selection that could produce this non-random pattern, strongly suggests that in some instances there are designed mechanisms driving mutations. The patterns seen here suggest that God, in His infinite wisdom, designed animals to be able to undergo genetic mutations which would enable them to adapt to a wide range of environmental challenges while minimizing risk.
Glossary | |
|---|---|
| Autosome: | a chromosome that is not a sex (X or Y) chromosome. |
| Centric fusion: | combining of two acrocentric (centromere near one end) chromosomes to form a new chromosome with the centromeres adjacent to each other. See figure 1. |
| Centromere transposition: | a change in the position of the centromere on the chromosome without a change in gene order. This rearrangement can be very difficult to distinguish from a pericentric inversion. |
| Frameshift: | an insertion or deletion (indel) that shifts the three-base-pair reading frame of the gene. A frameshift will often result in loss of function of the protein. |
| Haplotype: | a region of DNA usually inherited together; a group of alleles that are closely linked. |
| Heterochromatin: | sections of DNA containing highly repetitive sequences and few genes. Despite appearing inactive, these regions are important for proper function. The amount of heterochromatin appears to be adjusted following chromosomal rearrangements. |
| Karyotype: | the appearance of the chromosomes within an individual at metaphase, the time during cell division when the chromosomes are clearly seen. |
| Open reading frame: | the portion of DNA that is read (copied into RNA) and may be used for protein formation. |
| Knock-out mice: | mice in which the specific gene under study is disabled (knocked out). Studies with knock-out mice have been very helpful in determining the function of genes. |
| Paracentric inversion: | an inversion in one chromosome arm that does not include the centromere. See figure 1. |
| Pericentric inversion: | an inversion in a chromosome that includes the centromere. See Figure 1. |
| Phylogenomic: | comparison of the genomes of organisms within a group to attempt to reconstruct ancestry. |
| Single nucleotide polymorphism (SNP): | a difference in a single base in the DNA sequence; a change in which a single base pair differs from the usual base pair in that position. |
| Tandem fusion: | combining of two chromosomes where the end of one chromosome attaches to the end or centromeric region of another chromosome. See figure 1. |
| Tandem repeats: | multiple copies of the same base sequence on a chromosome. See figure 2. |
Related articles
Further reading
References
- Sanford, J.C., Genetic Entropy & The Mystery of the Genome, Elim Publishing, New York, 2005. Return to text.
- See Lightner, J.K., Designed to inhabit the earth, A review of The Plausibility of Life: Resolving Darwin’s Dilemma, Journal of Creation 22(1):33–36, 2008, and Williams, A., Facilitated variation: A new paradigm emerges in biology, Journal of Creation 22(1):85–92, 2008. Return to text.
- Siegler, H.R., The magnificence of kinds as demonstrated by canids, CRSQ 11(2):94–97, 1974. See also Wood, T.C., The current status of baraminology, CRSQ 43(3):149–158, 2006. Return to text.
- Genesis 7:1–10; Leviticus 11:27 Return to text.
- Ussher, J., The Annals of the World, Translated and edited by L. and M. Pierce, Master Books, Green Forest, AR, p. 19, 2003. Return to text.
- Fahey, B. and Myers, P., ‘Canidae’ (Online), Animal Diversity Web, 2000, animaldiversity.ummz.umich.edu/site/accounts/information/Canidae.html, accessed August 30, 2008. Return to text.
- Graphodatsky, A.S. et al., Phylogenomics of the dog and fox family (Canidae, Carnivora) revealed by chromosome painting, Chromosome Research 16(1):129–143, 2008. Return to text.
- Szczerbal, I. et al., Development of a cytogenetic map for the Chinese raccoon dog (Nyctereutes procyonoides procyonoides) and the arctic fox (Alopex lagopus) genomes, using canine-derived microsatellite probes, Cytogenetic and Genome Research 102: 267–271, 2003. Return to text.
- Szczerbal, I. et al., FISH mapping of 10 canine BAC clones harbouring genes and microsatellites in the arctic fox and the Chinese raccoon dog genomes, Journal of Animal Breeding and Genetics 123(5):337–342, 2006. Return to text.
- Lightner, J.K., Identification of species within the sheep-goat kind (Tsoan monobaramin), Journal of Creation 20(3):61–65, 2006. Return to text.
- Nachman, M.W. and Myers P., Exceptional chromosomal mutations in a rodent population are not strongly underdominant, Proceedings of the National Academy of Sciences of the United States of America 86(17):6666–6670, 1989. Return to text.
- Lightner, J.K., Changing chromosome numbers, Journal of Creation 20(3):14–15, 2006. Return to text.
- Lightner, J.K., Karyotype variability within the cattle monobaramin, Answers Research Journal 1:77–88, 2008. Return to text.
- ReMine, W., More precise calculations of the cost of substitution, CRSQ 43(2):111–120, 2006. Return to text.
- Rausch, V.R. and Rausch, R.L., Karyotype of the Red Fox, Vulpes vulpes L., in Alaska, Northwest Science 53(1):54–57, 1979. Return to text.
- Yudkin, D.V. et al., Mapping of KIT adjacent sequences on canid autosomes and B chromosomes, Cytogenetic and Genome Research 116:100–103, 2007. Return to text.
- Kennedy, L.J. et al., Association of a common dog leukocyte antigen class II haplotype with canine primary immune-mediated haemolytic anaemia, Tissue Antigens 68:502–508, 2006. Return to text.
- Angles, J.M. et al., Frequency and distribution of alleles of canine MHC-II DLA-DQB1, DLA-DQA1 and DLA-DRB1 in 25 representative American Kennel Club breeds, Tissue Antigens 66:173–184, 2005. Return to text.
- Kennedy et al., ref. 17. Return to text.
- Ito, H. et al., Allele frequency distribution of the canine dopamine receptor D4 gene exon III and I in 23 breeds, Journal of Veterinary Medical Science 66(7):815–820, 2004. Return to text.
- Hejjas, K. et al., Novel repeat polymorphisms of the dopaminergic neurotransmitter genes among dogs and wolves, Mammalian Genome 18(12):871–879, 2007. Return to text.
- Nöthen, M.M. et al., Human dopamine D4 receptor gene: frequent occurrence of a null allele and observation of homozygosity, Human Molecular Genetics 3(12):2207–2212, 1994. Return to text.
- Hejjas, K. et al., Association of polymorphisms in the dopamine D4 receptor gene and the activity-impulsivity endophenoyte in dogs, Animal Genetics 38(6):629–633, 2007. Return to text.
- Larsen, S.A. et al., Identification and characterization of tandem repeats in exon III of dopamine receptor D4 (DRD4) genes from different mammalian species, DNA Cell Biol 24(12):795–804, 2005. Return to text.
- Inoue-Murayama, M. et al., Sequence Comparison of the Dopamine Receptor D4 Exon III Repetitive Region in Several Species of the Order Carnivora, Journal of Veterinary Medical Science 64(8): 747–749, 2002. Return to text.
- Quignon, P. et al., The dog and rat olfactory repertoires, Genome Biology 6(10):R83, 2005. Return to text.
- Benbernou, N. et al., Functional analysis of a subset of canine olfactory receptor genes, Journal of Heredity 98(5):500–505, 2007. Return to text.
- Tacher, S. et al., Olfactory receptor sequence polymorphism within and between breeds of dogs, Journal of Heredity 96(7):812–816, 2005. Return to text.
- This is not only true because some pseudogenes have previously been shown to have important non-protein coding function, but also because a human OR pseudogene containing multiple premature stop codons has recently been demonstrated to code for a functional protein. Lai, P.C. et al., An olfactory receptor pseudogene whose function emerged in humans: a case study in the evolution of structure-function in GPCRs, Journal of Structural and Functional Genomics, September 19, 2008. Return to text.
- McBride, C.S., Rapid evolution of smell and taste receptor genes during host specialization in Drosophia sechellia, Proceedings of the National Academy of Sciences of the United States of America 104(12):4996–5001, 2007. Return to text.